- 移动端


武汉佰乐博生物技术有限公司品牌商
3 年
手机商铺
- NaN
- 0.20000000000000018
- 0.20000000000000018
- 2.2
- 2.2

推荐产品

公司新闻/正文
肠道再生之钥:科学家锁定小分子半胱氨酸
117 人阅读发布时间:2025-11-27 09:32
2025年10月,麻省理工学院科赫研究所的Ömer H. Yilmaz团队在顶刊Nature上发表题为“Dietary cysteine enhances intestinal stemness via CD8+ T cell-derived IL-22”了一项颠覆性研究,首次揭示了膳食中的半胱氨酸(一种常见于肉类、蛋奶中的必需氨基酸),能够像“特洛伊木马”一样被肠道吸收,暗中“策反”肠道内的CD8αβ⁺ T细胞,使其释放关键信号分子IL-22,从而大幅提升肠道干细胞的再生超能力。 这项研究不仅解开了营养调控免疫与干细胞再生的谜题,更为治疗放疗、化疗所致的肠道损伤乃至延缓肠道衰老提供了全新的“食疗”策略。

肠道作为营养物质消化和吸收的主要场所,其上皮细胞由位于隐窝底部的LGR5⁺肠道干细胞持续更新维持。既往研究表明,长期热量限制、高脂饮食等宏观营养变化可调控ISC功能,但特定氨基酸如何影响ISC命运及再生能力尚不明确。此外,肠道免疫微环境,尤其是CD8⁺ T细胞和IL-22等细胞因子,在调节ISC功能中扮演重要角色,然而营养代谢如何通过特定免疫细胞亚群调控IL-22产生并影响ISC功能,仍是未解之谜。
1. 半胱氨酸显著增强ISC介导的肠道再生能力
为探究氨基酸对肠道干细胞标志物HMGCS2表达的影响,研究者系统评估了20种蛋白源性氨基酸。结果显示,半胱氨酸处理可显著诱导小肠隐窝中HMGCS2的表达。进一步构建半胱氨酸富集饮食模型(CysRD),发现尽管小鼠体重增长减缓、内脏脂肪减少,但肠道形态和基础增殖无显著变化。然而,在体外类器官形成实验中,CysRD喂养小鼠的隐窝形成类器官效率显著提高,提示半胱氨酸增强了ISC的再生潜能。
在体内辐射损伤模型中,CysRD喂养显著增加了LGR5⁺和OLFM4⁺ ISC数量及BrdU⁺增殖细胞,并利用LGR5谱系追踪模型证实半胱氨酸处理后ISC及其子代细胞数量显著增加。此外,FITC-葡聚糖渗透性实验显示CysRD组屏障功能改善,而凋亡水平无差异,表明半胱氨酸主要通过增强增殖而非抑制凋亡来促进再生。
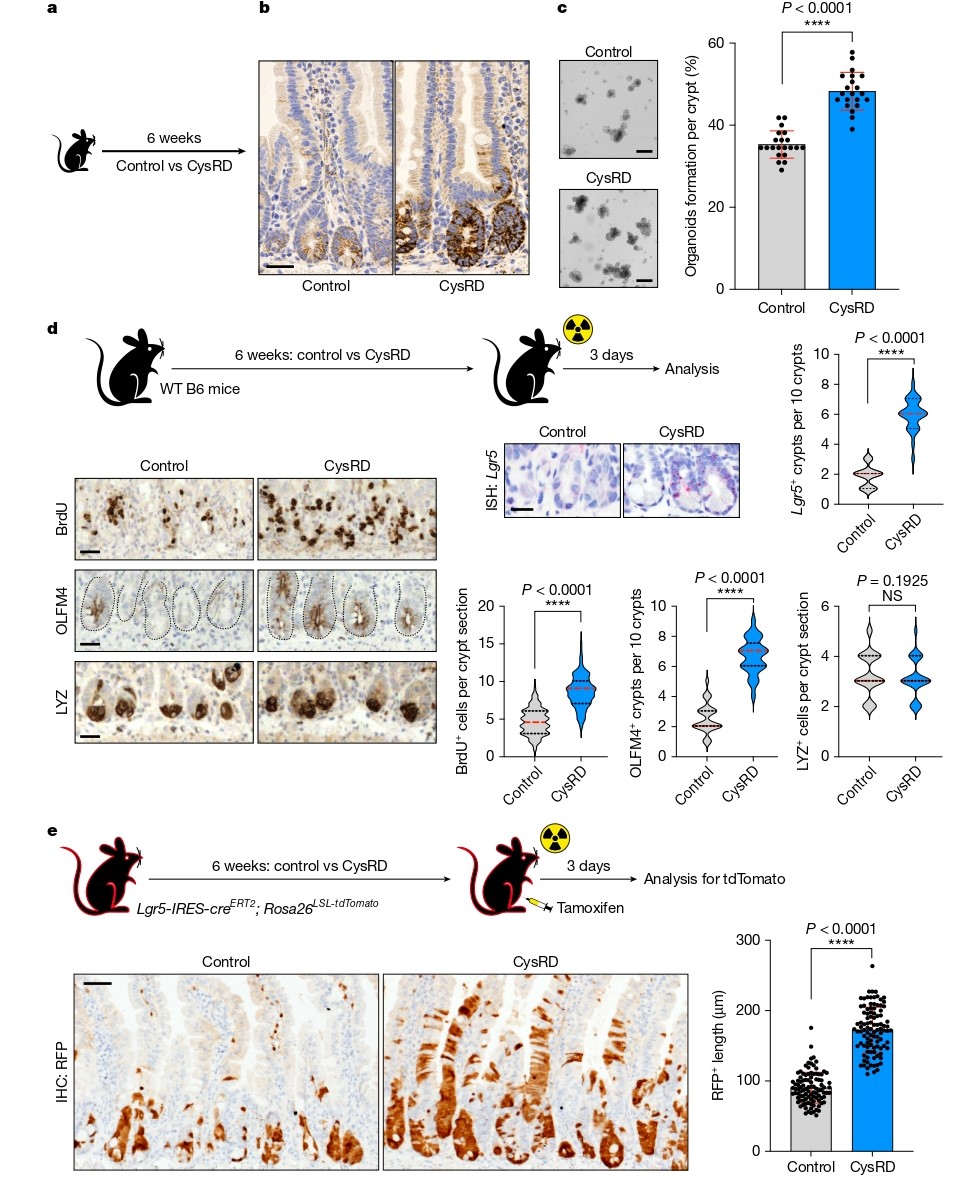
图1. 半胱氨酸增强肠道干细胞介导的再生功能
2. 半胱氨酸通过CD8αβ⁺ T细胞发挥促再生作用
研究者发现CysRD喂养显著增加了隐窝内CD8⁺ T细胞数量,尤其是CD8αβ⁺ T细胞亚群。单细胞RNA测序进一步证实CysRD组中效应型Gzmk⁺ CD8αβ⁺ T细胞比例上升,且TCR克隆分析显示其发生克隆扩增。机制上,CD8αβ⁺ T细胞的积累主要依赖于局部增殖增强,而非存活率改变或外源招募。
在Rag2⁻/⁻小鼠(缺乏成熟T、B细胞)中,半胱氨酸的促再生作用完全消失。共培养实验显示,CysRD来源的CD8αβ⁺ T细胞显著提升ISC类器官形成能力。更重要的是,抗体清除CD8αβ⁺ T细胞后,半胱氨酸驱动的肠道再生被显著抑制,明确了该细胞亚群在半胱氨酸作用中的必要性。
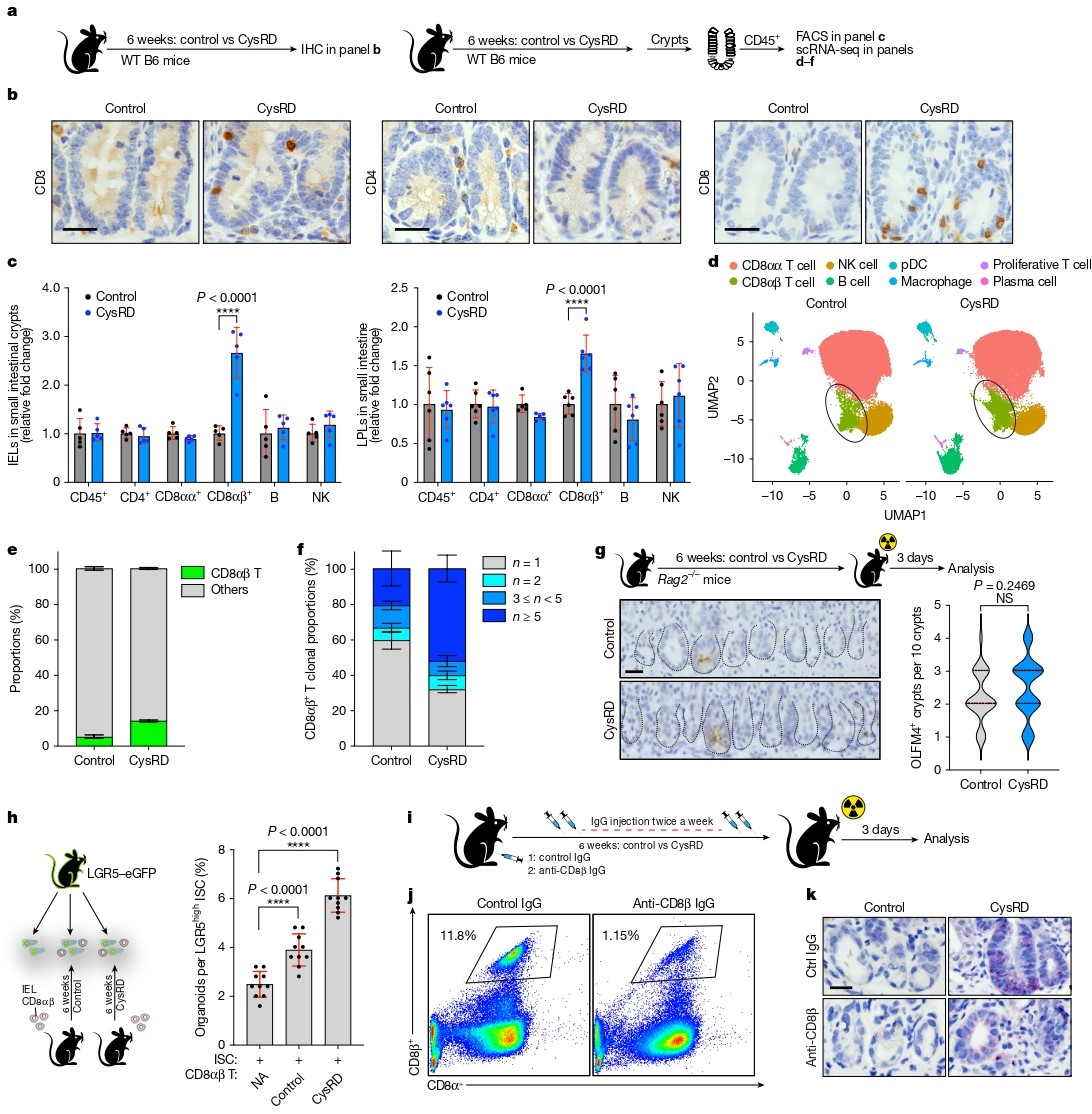
图2. 半胱氨酸驱动隐窝CD8αβ T细胞扩增并提升其增强干细胞功能的能力
3. CD8αβ⁺ T细胞来源的IL-22是关键效应因子
通过细胞因子阵列分析,研究者发现CysRD喂养后小肠中IL-22水平显著上升。进一步实验证实IL-22主要来源于CD8αβ⁺ T细胞,且外源性IL-22可诱导隐窝HMGCS2表达。在Il22⁻/⁻小鼠中,半胱氨酸虽仍能促进CD8αβ⁺ T细胞扩增,但无法诱导HMGCS2表达或增强ISC再生。
为验证CD8αβ⁺ T细胞来源的IL-22是否足以介导再生效应,研究者将野生型或Il22⁻/⁻ CD8αβ⁺ T细胞过继转移至Rag2⁻/⁻小鼠。结果显示,只有接受野生型T细胞的小鼠在CysRD喂养后表现出显著的ISC再生能力,明确IL-22是半胱氨酸–CD8αβ⁺ T细胞轴的下游关键效应因子。
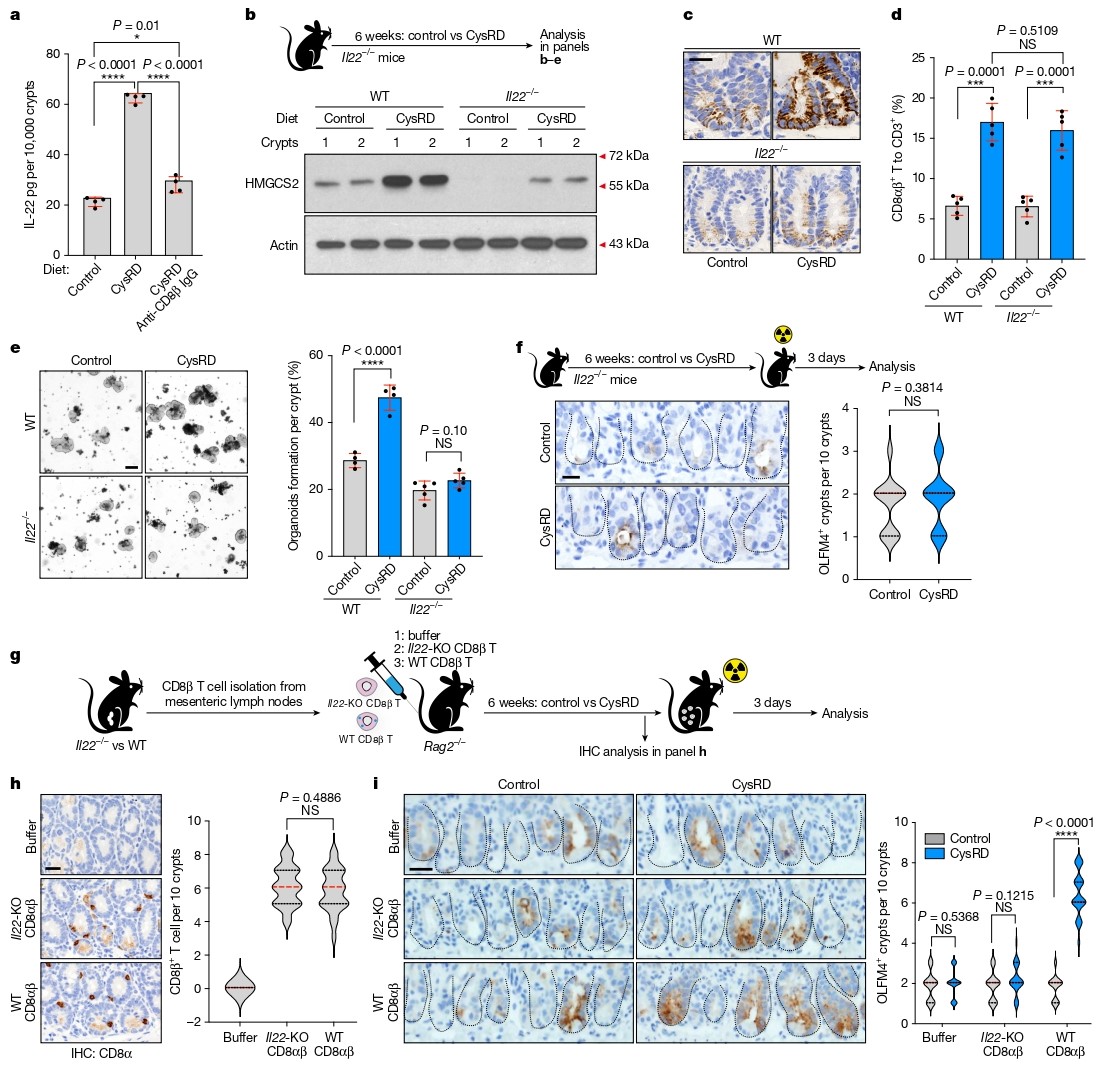
图3. 源自CD8αβ+ T细胞的IL-22介导半胱氨酸的促再生作用
4. 半胱氨酸代谢产物CoA驱动IL-22产生
代谢组学分析显示,CysRD喂养后小肠中半胱氨酸、胱氨酸及CoA水平显著上升。尽管维生素B5(泛酸)是CoA合成的另一前体,其补充未能复制半胱氨酸效应。而直接补充CoA可模拟半胱氨酸表型,包括诱导HMGCS2表达、促进CD8αβ⁺ T细胞扩增、增强IL-22产生及STAT3磷酸化,并改善辐射后肠道修复。体外实验中,CoA处理可直接诱导CD8αβ⁺ T细胞表达IL-22,明确了CoA在半胱氨酸–IL-22通路中的核心地位。
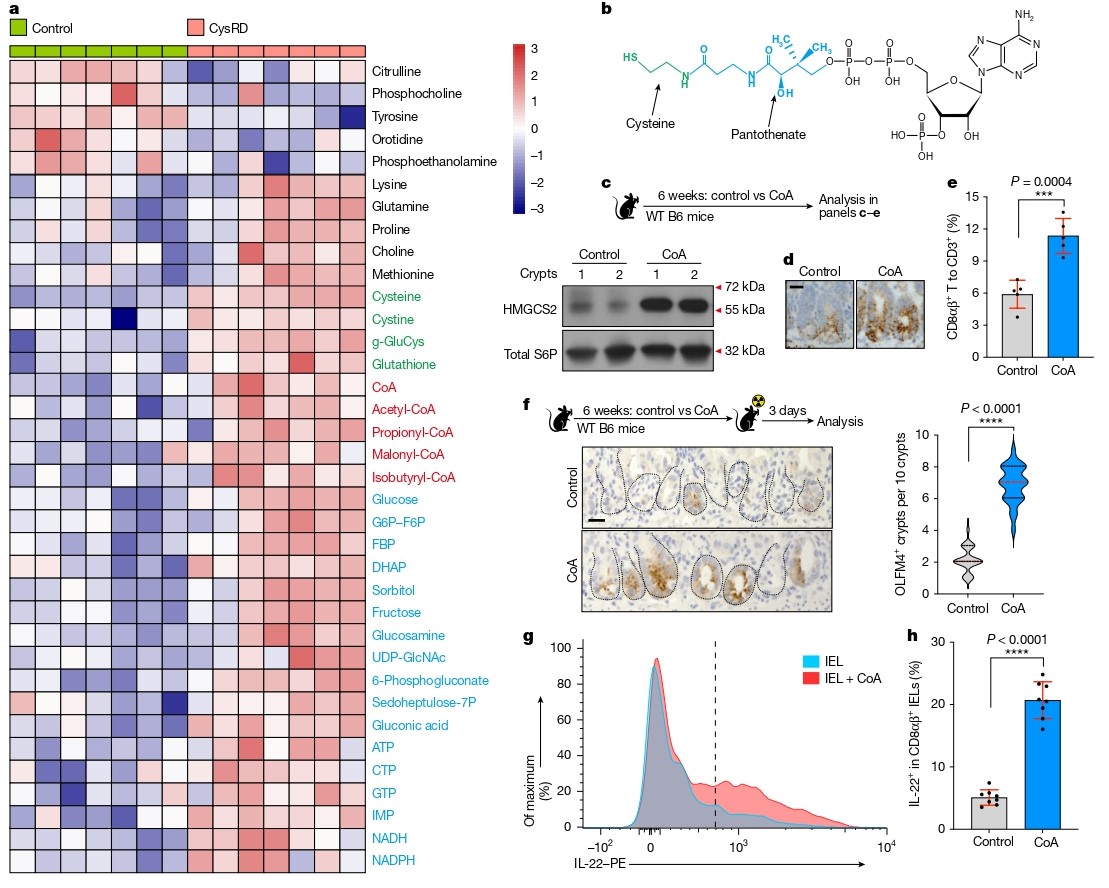
图4. 隐窝上皮细胞将半胱氨酸代谢为辅酶A,进而增强CD8αβ+ T细胞的IL-22生成
5. 上皮细胞胱氨酸摄取是启动再生信号的关键
为进一步解析细胞特异性机制,研究者构建了CD8⁺ T细胞特异性(*CD8-Cre; Slc7a11*ᶠ/ᶠ)和肠上皮特异性(Villin-Creᴱᴿᵀ²; Slc7a11ᶠ/ᶠ)Slc7a11敲除小鼠。结果显示,CD8⁺ T细胞自身缺失Slc7a11不影响半胱氨酸的促再生作用,而上皮特异性敲除则完全阻断了半胱氨酸诱导的HMGCS2表达、CD8αβ⁺ T细胞扩增、IL-22产生及组织修复。代谢组学分析进一步证实上皮Slc7a11缺失导致胱氨酸/半胱氨酸水平急剧下降,表明上皮细胞是 dietary cystine 摄取和下游信号启动的关键场所。
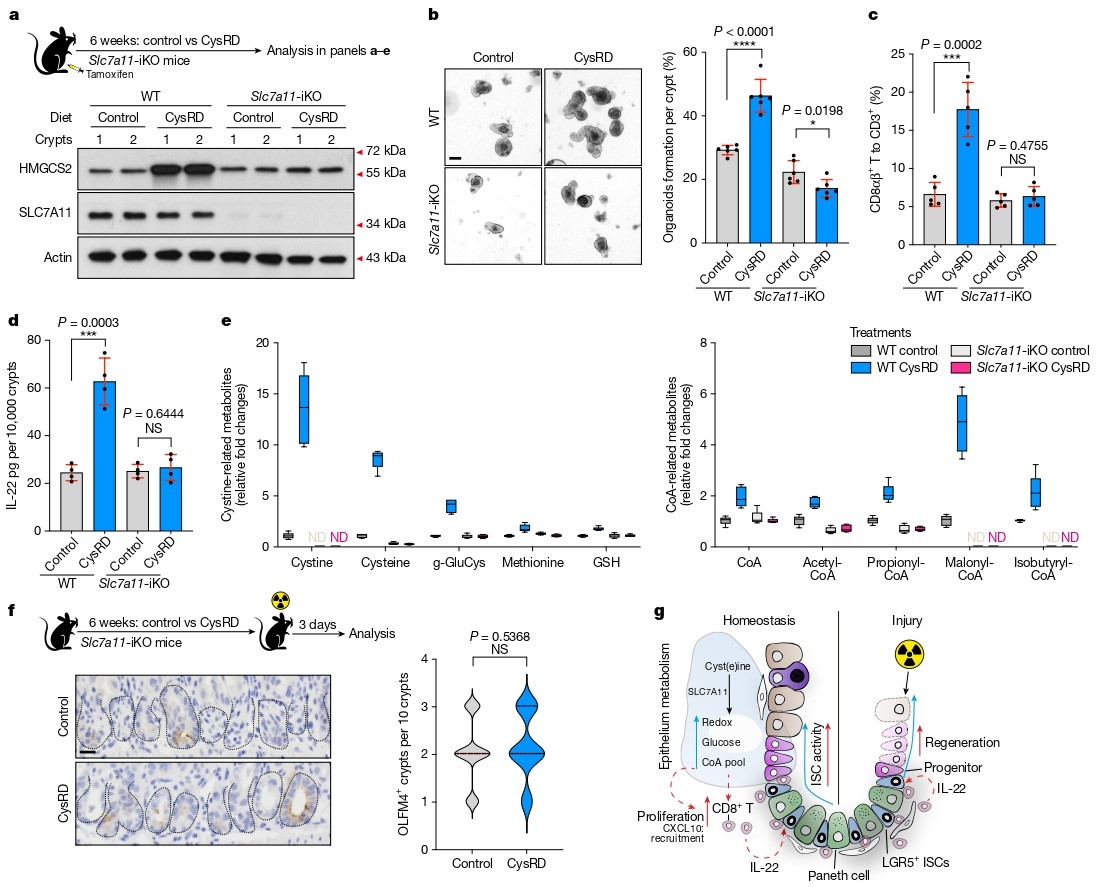
图5. 上皮细胞摄取半胱氨酸是辅酶A诱导CD8αβ T细胞产生IL-22、从而促进损伤后修复的关键
本研究首次揭示了“膳食半胱氨酸 → 上皮CoA合成 → CD8αβ⁺ T细胞扩增与IL-22产生 → ISC功能增强”这一跨细胞类型的代谢–免疫调控轴,深化了我们对营养–免疫–干细胞互作网络的理解。其创新性主要体现在三方面:
机制层面,研究明确了半胱氨酸并非通过其经典的抗氧化代谢物(如谷胱甘肽)发挥作用,而是通过CoA途径特异性调控CD8αβ⁺ T细胞的局部增殖与IL-22分泌。这一发现扩展了CoA在免疫代谢中的功能,提示其可能作为细胞间信号分子影响旁邻免疫细胞。
细胞互作层面,研究首次将CD8αβ⁺ T细胞确立为半胱氨酸调控ISC功能的关键中介,并揭示其通过IL-22直接增强ISC再生能力。这一发现不仅丰富了肠道免疫微环境的细胞图谱,也为理解特定T细胞亚群在组织修复中的作用提供了新视角。
区域特异性方面,研究阐明了半胱氨酸效应局限于小肠的机制:结肠中SLC7A11表达低、胱氨酸摄取能力弱,导致CoA合成与IL-22应答缺失。这一发现强调了营养感应与代谢应答的器官差异性,提示在营养干预中需考虑靶组织特性。
尽管该研究系统揭示了半胱氨酸的促再生机制,若干问题仍需深入探索。例如,CoA如何从上皮细胞释放并作用于CD8αβ⁺ T细胞?其转运机制尚未明确。此外,半胱氨酸是否在其他组织(如皮肤、造血系统)中通过类似机制调控干细胞功能?其在衰老或病理状态(如炎症性肠病)下的作用亦值得关注。
从转化视角看,半胱氨酸补充或CoA类似物可能成为增强肠道再生、改善化疗或放疗后黏膜损伤的潜在策略。尤其在与IL-22通路激动剂或其他免疫调节剂联用时,或可协同提升疗效。然而,半胱氨酸代谢在肿瘤发生中的作用复杂,其在肿瘤干细胞中的效应需审慎评估。
总而言之,该研究通过多学科技术手段,完整揭示了膳食半胱氨酸经由上皮代谢–免疫细胞互作–干细胞功能调控的跨细胞信号通路,为营养干预促进组织修复提供了新靶点与策略,也为理解代谢–免疫–干细胞轴在生理与病理过程中的作用奠定了重要基础。
AntibodySystem可提供本文使用的所有抗体,所有产品均经过严格验证,确保其在多种应用场景下的优异性能,助您在该突破性研究的基础上持续探索,推动您的肠道生物学与再生医学研究取得新突破。
|
货号 |
产品名称 |
|
RHJ09301 |
Anti-OLFM4 Antibody (R2H91) |
|
RGK32601 |
Anti-BrdU/5-BrdU Antibody (SAA0559) |
|
RHF45705 |
Anti-LYZ Antibody (R2A13) |
|
RXX54701 |
Anti-RFP/Red fluorescent protein Antibody (R1A04) |
|
RHE35909 |
Anti-CASP3/Caspase-3 Antibody (R3K78) |
|
FHK18512 |
Anti-Human CD8 Antibody (G10-1), PE |
|
RHF06801 |
Anti-HMGCS2 Antibody (R1Z03) |
|
FHD17430 |
Anti-Human EPCAM Antibody (C52) |
|
RHE27706 |
Anti-Phospho-STAT3 (Tyr705) Antibody (R3K42) |
|
RHE27704 |
Anti-STAT3 Antibody (R3K40) |
|
PHD57801 |
Anti-S6K1/RPS6KB1 Polyclonal Antibody |
|
RHK09202 |
Anti-SLC7A11/xCT Antibody (R2L62) |
|
VHC01901 |
InVivoMAb Anti-Human CXCL10/IP-10 (Iv0088) |
|
FHC34012 |
Anti-Human CD45/PTPRC Antibody (BC8), PE |
|
FHC27750 |
Anti-Human CD3 Antibody (SP34) |
|
PHB95901 |
Anti-CD4 Polyclonal Antibody |
|
RHD10805 |
Anti-CD19 Antibody (R3D98) |
|
FHG33810 |
Anti-Human CD161/KLRB1/NK1.1 Antibody (KM12.4.7) |
|
PHK17101 |
Anti-Human TRGV9/Vγ 9(TCR) Polyclonal Antibody |
|
RHD19402 |
Anti-CD127/IL7R Antibody (R3E63) |
|
RGK32601 |
Anti-BrdU/5-BrdU Antibody (SAA0559) |
|
FHJ53512 |
Anti-Human IL22 Antibody (SAA0391), PE |
佰乐博生物作为AntibodySystem亚洲独代,将为您的科研工作提供专业、全面、可靠的试剂产品。更多产品详情欢迎致电或者添加产品专员微信咨询!

扫码添加产品专员微信
电话:027-65279366
佰乐博官网:www.biolabreagent.com
咨询活性蛋白,活性抗体,请访问:
invivomab.cn
invivoplus.com